
لاکتات که مدتها تصور میشد صرفاً یک محصول جانبی متابولیسم بیهوازی و نشانهای از خستگی عضلانی است، به طور فزایندهای به عنوان یک مولکول سیگنالینگ حیاتی و یک سوبسترای انرژی همهکاره شناخته میشود. تحقیقات اخیر نقش پویای آن را در ارتباط بین اندامها و پردازش کربوهیدرات پس از غذا (postprandial) روشن کرده و پارادایمهای فیزیولوژیکی سنتی را به چالش میکشد. این مقاله دو مفهوم کلیدی را بررسی میکند که بر اهمیت لاکتات تأکید دارند: نظریه شاتل لاکتات و فرضیه شاتل لاکتات پس از غذا.
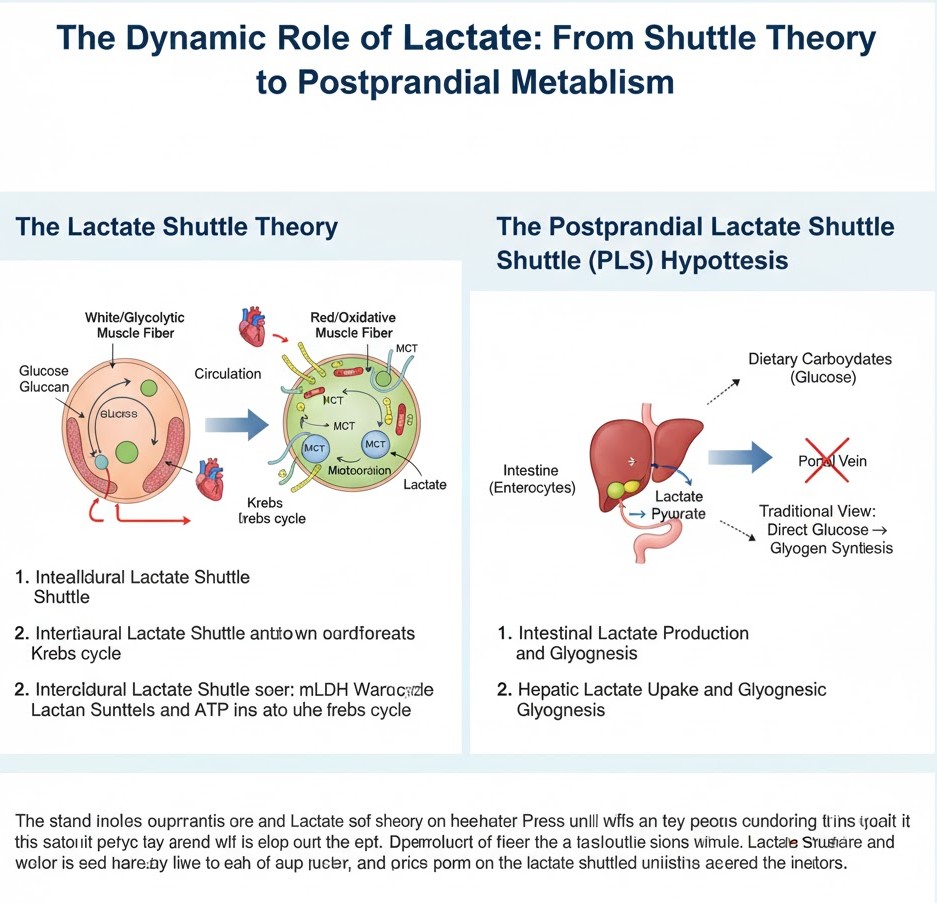
نظریه شاتل لاکتات: یک تغییر پارادایم در انتقال انرژی سلولی
نظریه شاتل لاکتات پیشنهاد میکند که لاکتات فقط یک زباله متابولیکی نیست، بلکه یک واسطه حیاتی در انتقال انرژی و ارتباط هم در داخل سلولها (به صورت داخل سلولی) و هم بین سلولها و بافتهای مختلف (به صورت بین سلولی) است.
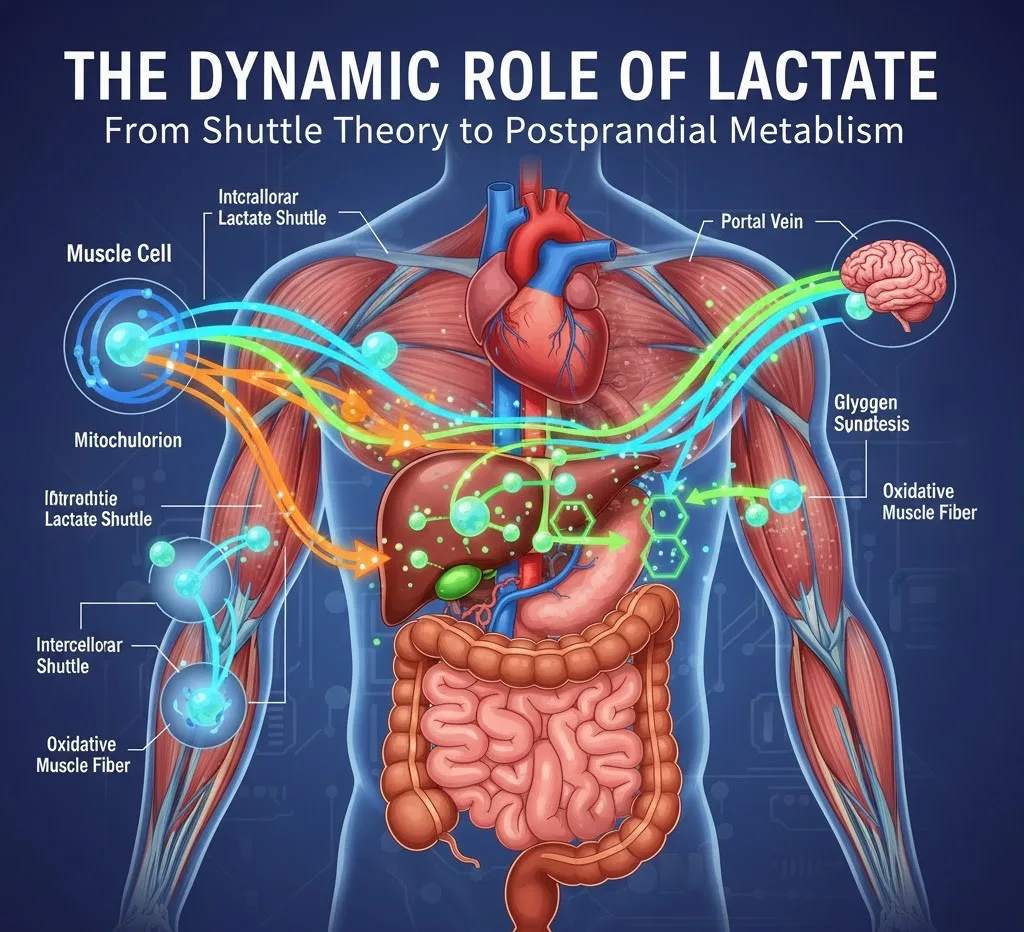
1. شاتل لاکتات داخل سلولی: در داخل یک سلول منفرد، لاکتات تولید شده در سیتوپلاسم (به عنوان مثال، در طی گلیکولیز) میتواند به داخل میتوکندری منتقل شود. در اینجا، توسط لاکتات دهیدروژناز میتوکندریایی (mLDH) دوباره به پیروات تبدیل میشود و متعاقباً برای اکسیداسیون کامل وارد چرخه Krebs میشود و ATP تولید میکند. این "شاتل داخل سلولی" امکان استفاده کارآمد از لاکتات را به عنوان منبع انرژی در همان سلولی که آن را تولید کرده است، فراهم میکند.
2. شاتل لاکتات بین سلولی: این جنبه از نظریه، حرکت لاکتات بین انواع سلولی یا اندامهای مختلف را توصیف میکند. سلولهایی با نرخ گلیکولیتیک بالا (به عنوان مثال، فیبرهای عضلانی تند انقباض، گلبولهای قرمز خون، برخی از سلولهای سرطانی) مقادیر قابل توجهی لاکتات تولید میکنند. این لاکتات سپس میتواند در جریان خون آزاد شود و توسط سلولهای دیگر (به عنوان مثال، فیبرهای عضلانی اکسیداتیو، قلب، مغز، کبد) که ظرفیت اکسیداتیو بالاتری دارند، جذب شود. این سلولهای "مصرف کننده" از لاکتات به عنوان یک منبع سوخت ترجیحی، به ویژه در دورههای تقاضای انرژی بالا، استفاده میکنند. انتقال دهندههای مونوکربوکسیلات (MCTs) نقش محوری در تسهیل این حرکت در سراسر غشای سلولی دارند.
نظریه شاتل لاکتات، لاکتات را به عنوان یک مولکول سیگنالینگ محوری، که هماهنگی متابولیکی را در سراسر بافتها و اندامهای مختلف سازماندهی میکند، تعریف میکند، نه صرفاً یک بن بست متابولیکی.
فرضیه شاتل لاکتات پس از غذا (PLS): یک دیدگاه جدید از متابولیسم کربوهیدرات
در حالی که نظریه شاتل لاکتات بر نقشهای عمومی لاکتات تمرکز دارد، فرضیه شاتل لاکتات پس از غذا (PLS) به طور خاص به نحوه پردازش کربوهیدراتهای غذایی پس از یک وعده غذایی میپردازد. این فرضیه یک دیدگاه جدید در مورد متابولیسم گلوکز، به ویژه در کبد ارائه میدهد.
به طور سنتی، اعتقاد بر این بود که پس از وعدههای غذایی غنی از کربوهیدرات، گلوکز جذب شده مستقیماً از طریق ورید پورت به کبد میرود، جایی که یا برای انرژی استفاده میشود یا به عنوان گلیکوژن ذخیره میشود. با این حال، فرضیه PLS یک مسیر ظریفتر را پیشنهاد میکند:
1. تولید لاکتات رودهای: بر اساس فرضیه PLS، بخش قابل توجهی از گلوکز غذایی ابتدا در داخل سلولهای رودهای (انتروسیتها) پس از جذب به لاکتات متابولیزه میشود. این فرآیند، که به عنوان "گلیکولیز هوازی" یا "اثر واربورگ" در روده شناخته میشود، ممکن است برای محافظت از کبد در برابر قرار گرفتن در معرض بیش از حد گلوکز و کمک به هموستاز انرژی روده عمل کند.
2. جذب لاکتات کبدی و گلیکوژنز: لاکتات تولید شده در روده سپس در گردش خون پورت آزاد میشود و ترجیحاً توسط کبد جذب میشود. در کبد، این لاکتات به عنوان یک سوبسترای اصلی برای گلوکونئوژنز (تولید گلوکز) یا، مهمتر از آن در این زمینه، برای سنتز گلیکوژن (گلیکوژنز) عمل میکند. این بدان معناست که مقدار قابل توجهی از گلیکوژن کبدی ممکن است از لاکتات سنتز شود تا مستقیماً از گلوکز.
پیامدها: فرضیه PLS درک مرسوم از تبدیل مستقیم گلوکز به گلیکوژن در کبد را به چالش میکشد. این فرضیه یک مسیر غیرمستقیم را پیشنهاد میکند که در آن روده به عنوان یک فیلتر متابولیکی اولیه عمل میکند و گلوکز را به لاکتات تبدیل میکند، که سپس به یک پیش ساز کلیدی برای گلیکوژن کبد تبدیل میشود. این میتواند پیامدهای قابل توجهی برای درک اختلالات متابولیکی، مانند دیابت نوع 2، و برای بهینه سازی استراتژیهای غذایی داشته باشد.
در نتیجه، هم نظریه شاتل لاکتات و هم فرضیه شاتل لاکتات پس از غذا، بر نقش چند وجهی و حیاتی لاکتات در فیزیولوژی انسان تأکید میکنند. آنها تصویری از لاکتات را نه به عنوان یک شرور متابولیکی، بلکه به عنوان یک واسطه پویا که توزیع انرژی، ارتباط بین اندامها و استفاده کارآمد از کربوهیدرات را تسهیل میکند، ترسیم میکنند و درک ما از تنظیم متابولیک را متحول میکنند.
در اینجا تصویری وجود دارد که مفاهیم را خلاصه میکند: